The mechanisms of cell-cell fusion
Cell-cell fusion is central for fertilization during sexual reproduction. It also underlies the formation of some tissues, such as muscles, during development. We study how haploid yeast gametes fuse to produce the diploid zygote during mating.
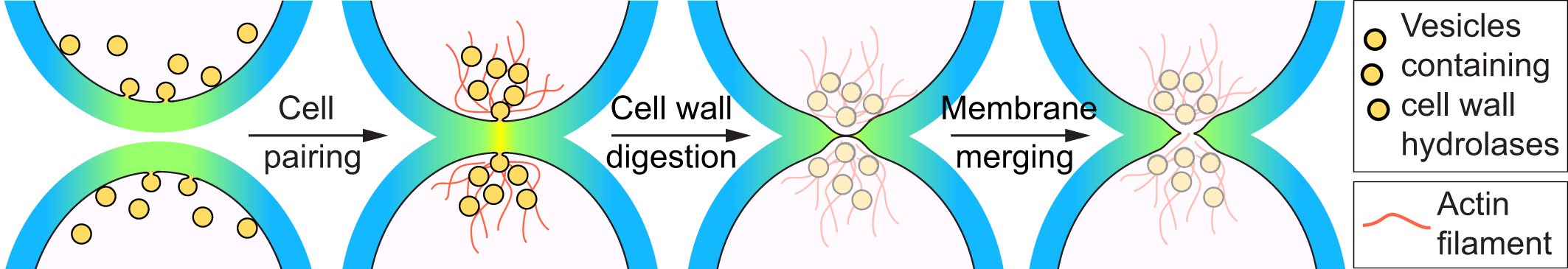
The fusion process starts after partner cells of distinct mating types have paired together, a process that relies on pheromone signaling, cell polarization and actin cytoskeleton re-organization. The fusion process can be conceptually divided in three stages. First, because fungal cells are encased in a rigid cell wall, the cells need to digest their cell wall at the zone of contact, leading to plasma membrane apposition. Second, the plasma membranes merge together to unite the two cells in one. Finally, the now diploid cell needs to establish its zygotic state and block further cell fusion attempts.
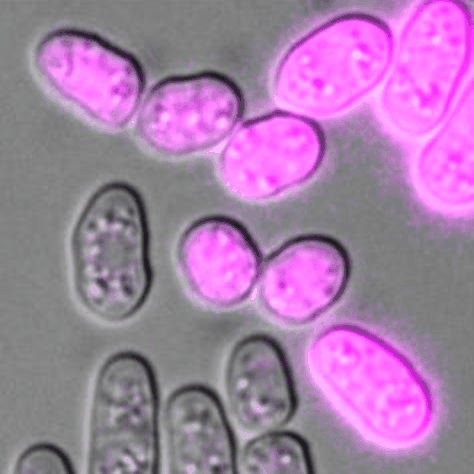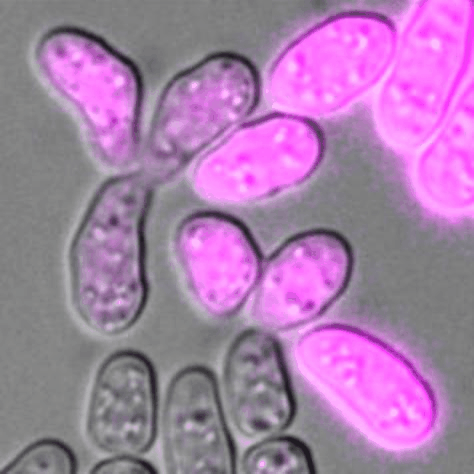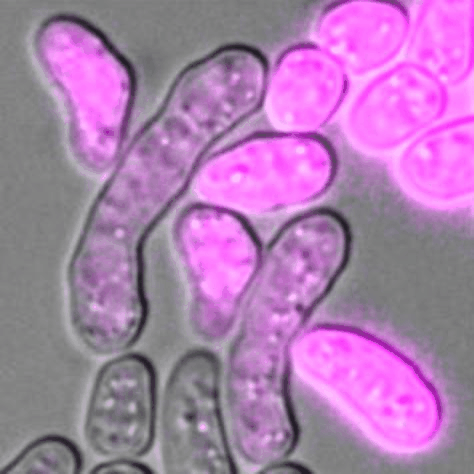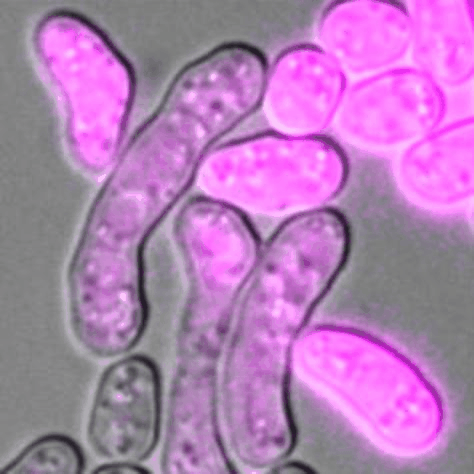 Cells of P (pink) and M (unlabelled) mating types during sexual reproduction
Cells of P (pink) and M (unlabelled) mating types during sexual reproduction
The actin fusion focus
Because the yeast cell wall offers protection not only against external insults but also against lysis due to high internal turgor pressure, the time and location of wall digestion has to be well controlled. Re-organization of the actin cytoskeleton is critical for local cell wall digestion. Fission yeast cells organize a specific actin structure – the actin fusion focus, an aster-like organization of actin filaments nucleated by the formin Fus1, which allows the focal accumulation of type V myosins. The myosins transport vesicles containing glucanases, enzymes that digest the cell wall. By contrast, cell wall synthases, which catalyze the formation of bonds between glucan polymers, are more broadly distributed at the cell-cell contact site. We propose that this geometric separation of cell wall hydrolases and synthases achieves the local cell wall digestion required for successful cell-cell fusion.
We study how the fusion focus is established. Its condensation relies on Fus1 formin self-interaction as well as on feedback between the formin and the myosin. It is also a site of pheromone communication and local signaling. Local pheromone-MAPK activation promotes focus stabilization. Because MAPK activation occurs in response to the partner’s pheromone, focus stabilization occurs only in paired cells. Negative regulation ensures that stabilization does not occur before cell pairing, which would lead to cell lysis.
Plasma membrane merging
The merging of the two cells’ plasma membrane is arguably the defining point in cell fusion. While a number of fusogenic proteins, which directly promote cell-cell fusion, are starting to emerge in various organisms, the fungal fusogen(s) remain enigmatic. We aim to discover the mechanisms of plasma membrane merging in yeast.
Preventing re-fertilization
How do zygotes prevent engaging additional gametes? This is critical to ensure genome maintenance across generations. We found that the fusion of P and M gametes leads to the rapid reconstitution of a bipartite transcription factor in the zygote, which blocks re-fertilization. This transcription factor consists of a M-cell-specific peptide Mi and a nuclear P-cell-specific homeobox protein Pi, which captures Mi in the P nucleus post-fusion. This asymmetric design leads to inherent asymmetric zygotic gene expression, which initiates first the Pi-containing nucleus. Delaying zygotic transcription leads to zygote fusing with additional gametes, thus forming polyploids and eventually aneuploid progeny.
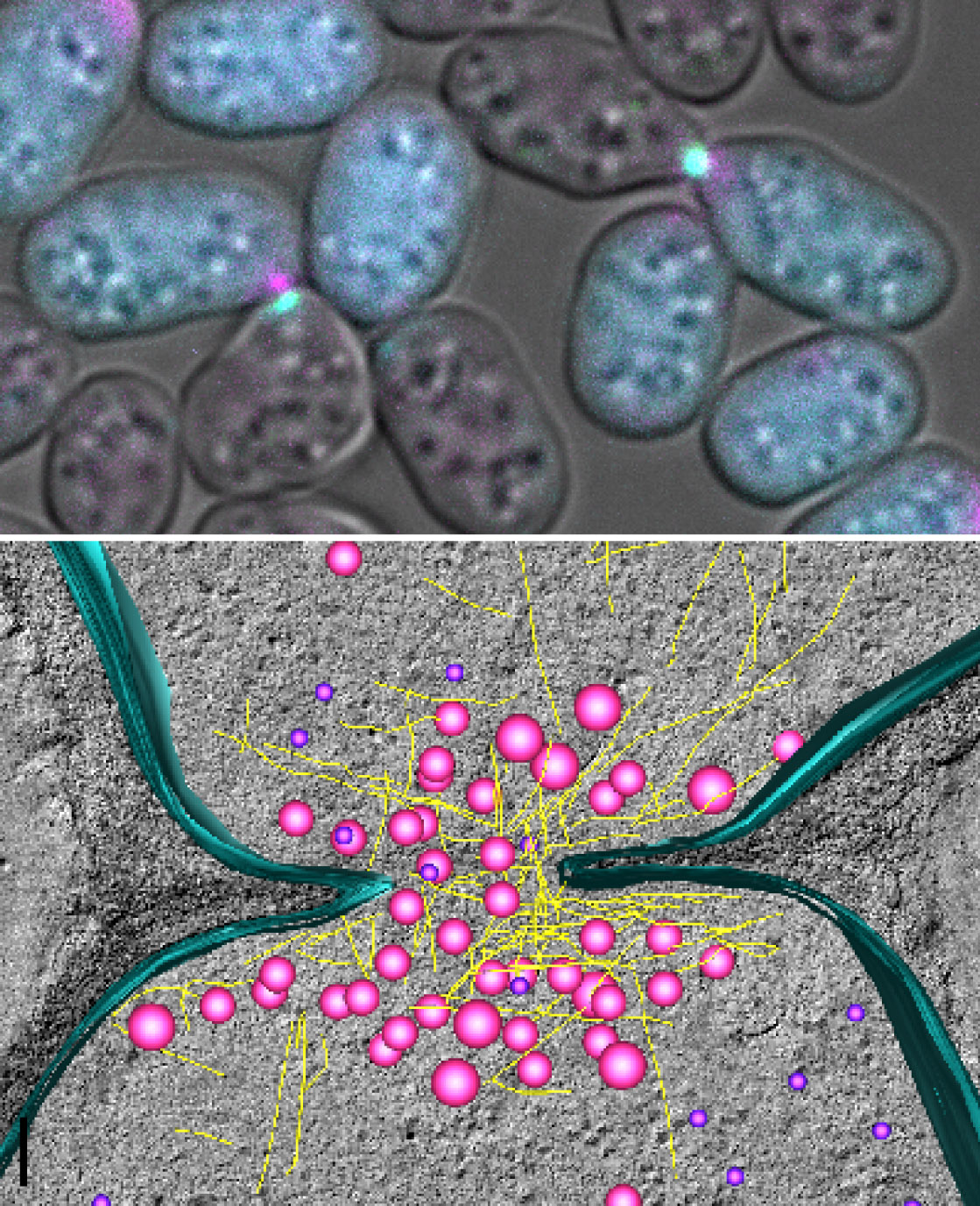
Imaging of mating cells by fluorescence microscopy (top) and electron microscopy (bottom)